Phylogeny of amphibians


Phylogeny of amphibians




Back to the
“MOLECULAR PHYLOGENETICS” page
Back to the
“LANE”
introduction page

FACTS
Species:
‣Ranid frogs
Locations:
‣All over the world, but mostly in India, Sri-Lanka, and Madagascar.

PUBLICATIONS
‣Journal of Evolutionary Biology, 7: 227-245 (1994)
‣Systematic Biology, 46: 126-144 (1997)
‣PNAS 97: 6585-6590 (2000)
‣Science 292: 93-95 (2001)
‣Science, 298: 379 (2002)
‣Science, 306: 479-481 (2004)
‣Science, 308: 199 (2005)
‣Systematic Biology 55(4): 579)-594, 2006

Related publications from our group on the phylogeny of other groups
‣Check the main “Molecular Phylogenetics” page

PEOPLE INVOLVED
FROM MICHEL
MILINKOVITCH’S LAB
‣Franky Bossuyt
Franky did his PhD in Milinkovitch’s lab on the phylogeny and phylogeography of Ranid frogs. He is now group leader at the Vrije Universiteit Brussel where he developed a research lab on amphibian evolution (web page)

MAIN COLLABORATORS
David Hillis &
David Cannatella
Both are Professors at the University of Texas, Austin, USA.
Jeff Powell
Professor at Yale University.
Gisella Caccone
Group leader at Yale University.




From European newts to Ranid frogs worldwide
While the use of molecular clocks was still in its infancy, one of my first molecular analyses (under the guidance of Gisella Caccone & Jeffrey Powell at Yale University) was to characterize the mitochondrial molecular clock in Euproctus newts using the disjunction and rotation of the Corsica-Sardinia microplate (from the Iberian Peninsula) as a calibration point.
My investigations on amphibian phylogenies started again with Franky Bossuyt, a bright student who did his PhD in my lab, starting in 1999. Franky is now group leader at the Vrije Universiteit Brussel where he developed a research lab on amphibian evolution (check his web page here). We focused on the phylogeny and phylogeography of ranid frogs ... a group that gave us some spectacular surprises (see below).
European newts
✓Caccone A., Milinkovitch M. C., Sbordoni V. & J. R. Powell
Molecular Biogeography: Using the Corsica-Sardinia Microplate Disjunction to Calibrate Mitochondrial rDNA Evolutionary Rates in Mountain Newts (Euproctus)
Journal of Evolutionary Biology, 7: 227-245 (1994)
✓Caccone A., Milinkovitch M. C., Sbordoni V. & J. R. Powell
Mitochondrial DNA Rates and Biogeography in European Newts (Genus Euproctus)
Systematic Biology, 46: 126-144 (1997)
➡ABSTRACT. Sequence divergence for segments of three mitochondrial DNA (mtDNA) genes encoding the 12s and 16s ribosomal RNA and cytochrome b was examined in newts belonging to the genus Euproctus (E. asper, E. montanus, E. platycephalus) and in three other species belonging to the same family (Salamandridae), Triturus carnifex, T. vulgaris, and Pleurodeles waltl. The three Euproctus species occur (one species each) in Corsica, Sardinia, and the Pyrenees. This vicariant distribution is believed to have been determined by the disjunction and rotation of the Sardinia-Corsica microplate from the Pyreneean region. Because time estimates are available for the tectonic events that led to the separation of the three landmasses, we used sequence data to estimate rates of evolution for the three gene fragments and investigated whether they conform to the rate-constancy hypothesis. By the Tajima (1993, Genetics 135:599-607) test, we could not detect rate heterogeneities for the ribosomal genes and for transversions in the cytochrome b gene. Assuming that these sites are evolving linearly over time and that cessation of gene flow occurred simultaneously with vicariant events, we compared the time of divergence estimated by molecular distances with the divergence times based on the geological estimates. Because we have two estimates of divergence time from the geological record, the split of Corsica/Sardinia from the Pyrenees and the split of Corsica from Sardinia, we could compare ratios of molecular divergence with the ratio of geological time divergence. The ratios are very similar, indicating that the molecular clock hypothesis cannot be rejected. These geological events also allowed us to calculate absolute rates of evolution for ribosomal and cytochrome b genes and compare them to rates for the same regions in other salamandrids and other vertebrates. Ribosomal mtDNA rates are comparable to those reported for other vertebrates, but cytochrome b rates are 3-7 times lower in salamanders than in other ectotherms. From a phylogenetic perspective, our data suggest that the cladogenic events leading to species formation in Euproctus and Triturus occurred very closely in time, indicating that the two genera may not be monophyletic. A duplication of the cytochrome b gene in T. carnifex was found, and the implications of this finding for mtDNA phylogenetic studies are discussed.
➡REMARK. When calibration of molecular clocks was in its infancy. Well, we were not too far off after all ;-)
Ranid frogs (and others)
With more than 1000 extant species, Ranidae sensu lato is one of the largest extant families of anurans. This cosmopolitan group of frogs evolved an enormous diversity in all kinds of habitats, such as forests, savannas, grasslands, and deserts. The characters associated with this ecological, morphological, and developmental diversity have been utilized extensively by systematists for generating hypotheses on ranid relationships. In recent years, the integration of molecular phylogenetic studies (e.g., see our refs below) led to revised classifications, recognizing several subfamilies with a wide distribution. However, because most investigations were based on relatively short gene sequences, or were limited to taxonomically or geographically restricted groups, a hypothesis for all major lineages of Ranidae is still lacking. In this study, we provide the first comprehensive estimate of the phylogenetic relationships of ranid frogs, with the secondary goal of evaluating concordance between evolutionary relationships and affinities suggested by current taxonomies. We used species sampled extensively from all currently and previously recognized taxonomic groups, and from all the continents. Second, we applied several combinations of calibration points, and used the union of these individual estimates as confidence intervals to obtain a conservative approximation of the timing of the major ranid diversifications. Third, we sought to evaluate the early biogeographic history of ranid frogs by evaluating our tree topologies and divergence time estimates in light of prevailing paleogeographic
models.

✓Bossuyt F. & M. C. Milinkovitch
Convergent Adaptive Radiations in Madagascan and Asian Ranid Frogs Reveal Co-variation between Larval and Adult Traits
PNAS 97: 6585-6590 (2000)
➡ABSTRACT. Recent studies have reported that independent adaptive radiations can lead to identical ecomorphs. Our phylogenetic analyses of nuclear and mitochondrial DNA sequences here indicate that a major radiation of ranid frogs on Madagascar produced morphological, physiological, and developmental characters that are remarkably similar to those that independently evolved on the Indian subcontinent. We demonstrate further that, in several cases, adult and larval stages each evolved sets of characters which are not only convergent between independent lineages, but also allowed both developmental stages to invade the same adaptive zone. It is likely that such covariations are produced by similar selective pressures on independent larval and adult characters rather than by genetic or functional linkage. We briefly discuss why larval/adult covariations might constitute an important evolutionary phenomenon in species for which more than one developmental stage potentially has access to multiple environmental conditions.
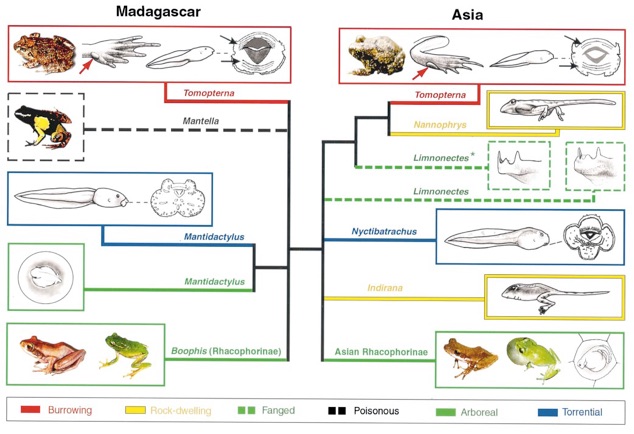
Figure: Convergences in larval and adult ecomorphs between and within Madagascan and Asian ranids. All Asian species are from the Indian subcontinent, except one (Limnonectes kuhlii, indicated by an asterisk) coming from Vietnam. Adult burrowing frogs (red boxes), beside having a toad-like general morphology and short hind limbs, exhibit a suite of characters [such as feet with a crescentic inner metatarsal tubercle (red arrows) and partly connected lateral metatarsals] that are adaptations to the fossorial zone. Their tadpoles have a general body shape and mouth parts [such as rows of keratinized teeth (black arrows)] which are of the typical ancestral aquatic type. Arboreal frogs (green boxes) exhibit similar adult ecomorphs in Madagascan and Indian Rhacophorinae. Some Asian tree frogs exhibit a development on land with complete metamorphosis in the egg, showing a remarkable convergence with some Madagascan arboreal species (Mantidactylus). Some rock-dwelling frogs (yellow boxes) have semi-terrestrial larvae with stout hind-limbs, a strongly developed tail with much reduced fin membranes, and with the ventral side of the body and the spiracle tube (as well as, sometimes, the anal tube) flattened. Certain torrential frogs (blue boxes) have larvae that lack horny teeth and exhibit, around the mouth, enlarged lobes that are richly provided with papillae. Fanged frogs (dashed green boxes) have protruding fangs in the lower jaw. Although poisonous frogs do not occur in Asia, some poisonous mantellas (black dashed box) exhibit aposematic colors and are remarkably convergent with the neotropical poison arrow frogs, Dendrobatidae.
➡REMARK. The number and amplitude of convergence events we uncovered in this study stroke me ever since. This is (together with my PNAS paper on the polyphyly of river dolphins) what made me interested into Evo-Devo. Obviously, we had to give up the idea of performing evo-devo studies on these taxa (river dolphins and Asian/Madagascan ranid frogs are not the easiest laboratory models for studying the molecular mechanisms that generate convergent traits). Hence, we switched to new laboratory model systems (see our Evo-Devo web pages).

✓Bossuyt F. & M. C. Milinkovitch
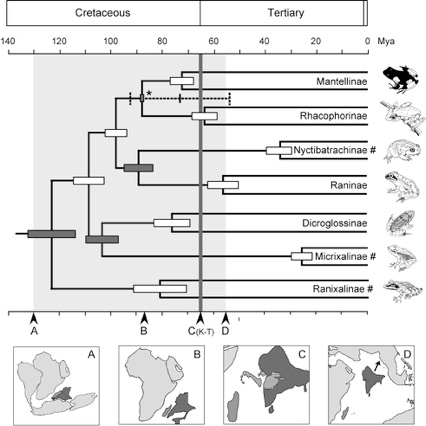
Amphibians as Indicators of Early Tertiary 'Out-of-India' Dispersal of Vertebrates
Science 292: 93-95 (2001)


✓Meegaskumbura M., Bossuyt F., Pethiyagoda R., Manamendra-Arachchi K., Bahir M.,
Milinkovitch M. C. & C. J. Schneider
Sri Lanka: A New Amphibian Hotspot
Science, 298: 379 (2002)
➡REMARK. There is no abstract as it is Science “Brevia”. In that paper, we report the discovery of an endemic radiation of >100 species of Old World tree frogs (Rhacophorinae) from Sri Lanka, an island from which only 18 rhacophorine species were known previously. An extensive survey, primarily in the island’s remaining 750 km2 of rain forest, revealed upwards of 140 anuran species, making Sri Lanka an amphibian hot spot of global importance.

✓Bossuyt F., Meegaskumbura M., Beenaerts N., Gower D.J., Pethiyagoda R., Roelants K., Mannaert A., Wilkinson M., Bahir M. M., Manamendra-Arachchi K., Ng P.K.L., Schneider C. J., Oommen O.V. & Michel C. Milinkovitch
Local Endemism Within the Western Ghats-Sri Lanka Biodiversity Hotspot
Science, 306: 479-481 (2004)
➡ABSTRACT. The apparent biotic affinities between the mainland and the island in the Western Ghats–Sri Lanka biodiversity hotspot have been interpreted as the result of frequent migrations during recent periods of low sea level. We show, using molecular phylogenies of two invertebrate and four vertebrate groups, that biotic interchange between these areas has been much more limited than hitherto assumed. Despite several extended periods of land connection during the past 500,000 years, Sri Lanka has maintained a fauna that is largely distinct from that of the Indian mainland. Future conservation programs for the subcontinent should take into account such patterns of local endemism at the finest scale at which they may occur.

➡Associated publication:
✓Bossuyt F., Meegaskumbura M., Beenaerts N., Gower D.J., Pethiyagoda R., Roelants K., Mannaert A., Wilkinson M., Bahir M.M., Manamendra-Arachchi K., Ng P.K.L., Schneider C. J., Oommen O.V. & M. C. Milinkovitch
Biodiversity in Sri Lanka and the Western Ghats
Science, 308: 199 (2005)

✓Bossuyt F., Brown R.M., Hillis D.M., Cannatella D.C. & M. C. Milinkovitch
Phylogeny and Biogeography of a Cosmopolitan Frog Radiation: Late Cretaceous Diversification Resulted in Continent-scale Endemism in the Family Ranidae
Systematic Biology 55(4): 579)-594, 2006
➡ABSTRACT. Ranidae is a large anuran group with a nearly cosmopolitan distribution. We investigated the phylogenetic relationshipsandearly biogeographic history of ranid frogs, using 104 representatives of all subfamiliesandfamilies, sampled from throughout their distribution. Analyses of about 1570 bp of nuclear gene fragments (Rag-1, rhod, Tyr) and about 2100 bp of the mitochondrial genome (12S rRNA, tRNAVAL , 16S rRNA) indicate that the monophyly of several taxa can be rejected with high confidence. Our tree is characterized by a clear historical association of each major clade with one Gondwanan plate. This prevalence of continent-scale endemism suggests that plate tectonics has played a major role in the distribution of ranid frogs. We performed dispersal-vicariance analyses, as well as analyses constrained by paleogeographic data, to estimate ancestral distributions during early ranid diversification. Additionally, we used molecular clock analyses to evaluate whether these scenarios fit the temporal framework of continental breakup. Our analyses suggest that a scenario in which the ancestors of several clades (Rhacophorinae, Dicroglossinae, Raninae) reached Eurasia via the Indian subcontinent, and the ancestor of Ceratobatrachinae entered via the Australia–New Guinea plate, best fits the paleogeographic models and requires the fewest number of dispersal/vicariance events. However, several alternatives, in which part of the ranid fauna colonized Laurasia from Africa, are not significantly worse. Most importantly, all hypotheses make clear predictions as to where to expect key fossils and where to sample other living ranids, and thus constitute a strong basis for further research.
➡REMARK. This is a large study putting our “Out-of-India” hypothesis into context with much additional paleogeographic models, and greatly extending the taxonomic sampling.

